
Dendroctone du pin ponderosa (fiche d'information)
Nous avons archivé cette page et elle ne sera plus mise à jour.
Vous pouvez la consulter à des fins de recherche ou à titre de référence.

Nom commun anglais : Mountain pine beetle
Nom scientifique : Dendroctonus ponderosae Hopkins
Ordre : Coléoptères
Famille : Curculionidés
Répartition
Le dendroctone du pin ponderosa est une espèce indigène de l’ouest de l’Amérique du Nord répartie du nord du Mexique au nord de la Colombie-Britannique. Il est également présent dans une population isolé de pin entourée par la prairie dans la région des collines du Cyprès, dans le sud-ouest de la Saskatchewan. Cette population a vraisemblablement été infestée au début des années 1980. Au cours des années 2000, le ravageur a considérablement étendu son aire de répartition au Canada, envahissant un nouvel habitat à l’est des Rocheuses, dans le nord-est de la Colombie-Britannique et le nord de l’Alberta.
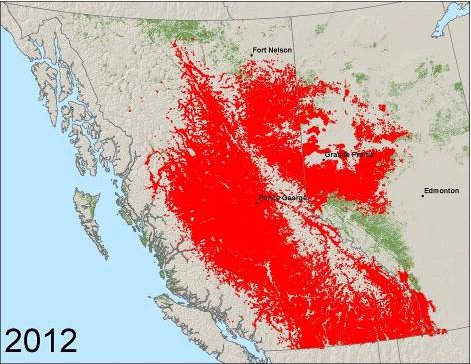
Présence observée du dendroctone du pin ponderosa de 1999 à 2012.
Données cartographiques : Forest Practices Branch, Ministry of Forests and Range, gouvernement de la Colombie-Britannique; Environment and Sustainable Resource Development, gouvernement de l’Alberta; Relevé des insectes et des maladies des arbres, Centre de foresterie du Pacifique, Service canadien des forêts, Ressources naturelles Canada.
Hôtes au Canada
Principaux hôtes au Canada
Le pin tordu est l’hôte de prédilection du dendroctone du pin ponderosa en Colombie-Britannique.
Autres hôtes
La plupart des espèces de pins qui poussent dans l’aire de répartition du ravageur sont fréquemment attaquées, à l’exception du pin de Jeffrey. Au Canada, le dendroctone du pin ponderosa attaque le pin ponderosa, le pin argenté, le pin à blanche écorce et le pin flexible et, depuis peu, le pin gris, dans les régions nouvellement colonisées.
Hôtes occasionnels ou potentiels
La plupart des espèces de pins indigènes au Canada, de même que quatre espèces non indigènes, ont été attaquées avec succès lorsqu’elles ont été plantées dans des régions comprises dans l’aire de répartition du ravageur dans l’ouest de l’Amérique du Nord. Certains conifères autres que les pins tels que l’épinette de l’intérieur, le douglas et le mélèze occidental sont parfois attaqués durant les infestations lorsqu’ils poussent avec des pins, mais les populations du dendroctone ne persistent pas longtemps dans de tels hôtes.
Cycle vital

Le dendroctone du pin ponderosa a un cycle d’un an dans la plupart des régions comprises dans son aire de répartition, mais il peut boucler son développement plus ou moins rapidement selon les températures locales. Selon la région, les adultes se dispersent habituellement en juillet ou en août pour coloniser de nouveaux hôtes. Les femelles sont les premières à attaquer et émettent des substances sémiochimiques appelées phéromones d’agrégation pour attirer d’autres femelles et les mâles vers l’arbre qu’elles ont colonisé. Elles déposent leurs œufs sur les parois des galeries verticales qu’elles forent dans l’écorce interne de l’hôte. Les larves néonates forent leur propre galerie perpendiculairement à la galerie parentale. Le dendroctone du pin ponderosa hiberne habituellement au stade larvaire, poursuivant son développement au cours du printemps suivant et se nymphosant en juin ou en juillet.
Durant le forage de leurs galeries, les dendroctones inoculent dans les tissus de l’hôte des champignons symbiotiques qu’ils transportent dans des poches spécialisées dans une de leurs pièces buccales. La colonisation de l’écorce interne et de l’aubier par les champignons entrave les fonctions et neutralise les mécanismes de défense de l’hôte et modifie les conditions d’humidité et la chimie des tissus de l’hôte dans lesquels les larves se développent. Les dendroctones ténéraux se nourrissent des spores libérées par les champignons dans leur loge nymphale avant d’émerger et de se disperser pour coloniser un nouvel hôte.
Écologie

Les arbres attaqués se défendent contre le ravageur en produisant de la résine toxique. Les populations de faible densité ou endémiques ne parviennent pas à surmonter les mécanismes de défense des arbres sains et attaquent les sujets supprimés, affaiblis ou mourants. Ces derniers sont habituellement des hôtes de piètre qualité parce qu’ils ont souvent déjà été attaqués par des insectes compétiteurs et que leur couche d’écorce interne, du fait de sa minceur, constitue elle-même un habitat de piètre qualité.
Lorsque les populations augmentent, le dendroctone du pin ponderosa parvient à surmonter les défenses des arbres de plus grande taille et en meilleure santé en menant des attaques massives rapides et coordonnées. Les grands arbres constituent un meilleur habitat et permettent le développement d’un plus grand nombre de dendroctones. Cela crée une boucle de rétroaction positive qui favorise la rapidité de croissance des populations. Les défenses des hôtes jouent peut-être un rôle important dans la régulation des populations de faible densité ou endémiques, mais elles sont inefficaces lorsque les populations atteignent des densités élevées. Les grandes infestations à l’échelle du paysage commencent à décliner seulement lorsque la plupart des hôtes de grand diamètre sont morts et que des conditions atmosphériques défavorables déciment les populations du ravageur.
En Colombie-Britannique, la limite nord de l’aire de répartition du dendroctone du pin ponderosa était limitée par les grands froids hivernaux (− 40 °C) et les étés frais. Les infestations ont été associées à la présence de conditions météorologiques favorables en été et en hiver. Les étés chauds et secs favorisent le développement et la dispersion du ravageur, et le stress dû à la sécheresse affaiblit les mécanismes de défense des arbres hôtes. La mortalité hivernale est habituellement la plus importante cause de mortalité chez le dendroctone du pin ponderosa; à l’inverse, les hivers doux ont un effet bénéfique sur la survie du ravageur. La présence de conditions météorologiques favorables au cours de plusieurs étés et hivers consécutifs et l’abondance de pins hôtes murs de qualité à l’échelle du paysage ont été citées comme deux facteurs ayant contribué au déclenchement des infestations massives qui se sont produites en Colombie-Britannique au cours des années 1990 et 2000.
Attaques et dommages

La sciure qui s’accumule sur l’écorce externe autour des trous d’entrée forés par les dendroctones est le premier signe d’attaque. Dans les jours qui suivent les premières attaques, des tubes de résine (petites gouttes de résine) apparaissent sur l’écorce externe des arbres infestés. Ces tubes demeurent visibles pendant des années, mais ils peuvent ne pas être présents chez les arbres dont les défenses sont gravement compromises. L’aubier des arbres attaqués avec succès prend une teinte bleutée, habituellement dès l’automne suivant l’attaque initiale. Les aiguilles virent au jaune, à l’orange puis au rouge environ un an après l’attaque initiale. La cime de certains arbres peut commencer à se décolorer dès la première année, selon les conditions environnementales. Les aiguilles roussies tombent environ deux à quatre ans après l’attaque initiale, et les arbres dénudés semblent gris.
Situation au Canada
La plus grave infestation documentée s’est produite au cours des années 1990 et 2000 en Colombie-Britannique. Plus de 18 millions d’hectares de forêt ont été touchés à des degrés divers, totalisant une perte d’environ 723 millions de mètres cubes (53 %) du volume marchand de pin en date de 2012. L’infestation a atteint son point culminant en 2005. On prévoit que les pertes cumulées totales atteindront 752 millions de mètres cubes (58 %) du volume marchand de pin en 2017. L’infestation se sera alors en grande partie résorbée en Colombie-Britannique.
Plusieurs fois au cours des années 2000, des adultes issus de l’infestation massive sévissant dans le centre de la Colombie-Britannique ont été emportés par des vents soufflant dans la haute atmosphère au-dessus de la barrière biogéoclimatique formée par les Rocheuses et déposés dans le nord-est de la Colombie-Britannique et le nord-ouest de l’Alberta. Ces incidents de dispersion sur de grandes distances ont contribué à accroître considérablement l’aire de répartition du ravageur au Canada. Le dendroctone du pin ponderosa est maintenant établi dans les forêts de pin tordu et de pin gris du nord de l’Alberta et risque de se disperser vers l’est à travers toute la forêt boréale canadienne si les conditions lui sont favorables. Il a également étendu son aire de répartition vers le nord, et en 2012, sa présence a été signalée pour la première fois au nord du 60e parallèle, dans les Territoires du Nord-Ouest. Le sort de cette petite population est toutefois incertain.

Photo : K. Buxton, BC Ministry of Forests, Lands and Natural Resource Operations de la Colombie-Britannique.
Contact : Kathy Bleiker
Liens
Publications du Service canadien des forêts sur le dendroctone du pin ponderosa
Références
Références
Cerezke, H.F., et F.J. Edmond. 1989. Forest insect and disease conditions in Alberta, Saskatchewan, Manitoba, and the Northwest Territories in 1987. Rapport d’information NOR-X-300. Edmonton, Alberta : Centre de foresterie du Nord, Service canadien des forêts, Ressources naturelles Canada.
Furniss, M.M., et J.A. Schenk. 1969. Sustained natural infestations by the mountain pine beetle in seven new Pinus and Picea hosts. Journal of Economic Entomology 62: 518–519.
Paine, T.D., K.F. Raffa, et T.C. Harrington. 1997. Interactions among scolytid bark beetles, their associated fungi, and live host conifers. Annual Review of Entomology 42: 179–206.
Raffa, K.F., B.H. Aukema, B.J. Bentz, A.L. Carroll, J.A. Hicke, M.G. Turner, et W.H. Romme. 2008. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. BioScience 58: 501–517.
Safranyik, L., et A.L. Carroll. 2006. The biology and epidemiology of the mountain pine beetle in lodgepole pine forests. Dans L. Safranyik et B. Wilson (éd.), The mountain pine beetle: A synthesis of its biology, management and impacts on lodgepole pine. Victoria, Colombie-Britannique : Centre de foresterie du Pacifique, Service canadien des forêts, Ressources naturelles Canada.
Safranyik, L., A.L. Carroll, J. Régnière, D.W. Langor, W.G. Riel, B. Peter, B.J. Cooke, V.G. Nealis, S.W. et Taylor. 2010. Potential for range expansion of mountain pine beetle into the boreal forest of North America. Canadian Entomologist 142: 415–442. doi:10.4039/n08-CPA01.
Smith, R.H., J.P. Cramer, et E.J. Carpender. 1981. New record of introduced hosts for the mountain pine beetle in California. Research Note PSW-354, United States Forest Service.
Walton, A. 2013. Provincial-level projection of the current mountain pine beetle outbreak: update of the infestation projection based on the Provincial Aerial Overview Surveys of Forest Health conducted from 1999 through 2012 and the BCMPB model (year 10). Victoria, Colombie-Britannique : Ministry of Forests, Lands and Natural Resources Operations de la Colombie-Britannique.
Wood, S.L. (éd.). 1982. The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae): a taxonomic monograph. Provo, Utah: Brigham Young University.
Détails de la page
- Date de modification :