
Applications en foresterie
Nous sommes au fait de la nécessité constante de comprendre et quantifier l'état et la dynamique des forêts aux échelles régionale et planétaire. Parmi les renseignements nécessaires, on retrouve la cartographie selon le type de forêts, la découverte des surfaces coupées à blanc et des incendies récents, et l'extraction de plusieurs données biophysiques, dont la biomasse totale et l'âge des arbres.
L'on prévoit que l'utilisation des données polarimétriques facilitera la détection des différences structurelles entre les couverts forestiers et, ainsi, contribuera à cartographier ces types de forêt et apportera des renseignements supplémentaires à d'autres applications de gestion de la forêt. La figure 9-26 montre une image composite en fausses couleurs de données multipolarimétriques d'une région à l'est d'Ottawa et illustre l'exploitation des informations additionnelles dans l'indentification forestière.
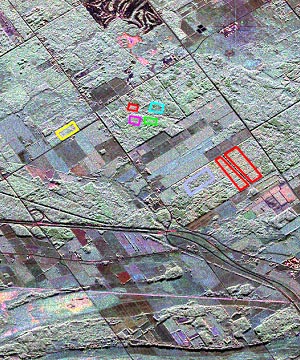
Figure 9-26. Image composite en fausses couleurs du site d'étude de la Mer bleue près d'Ottawa. Sont illustrées, six zones forestières peuplées de diverses essences (données RSO-C, rouge : HH, vert : HV, bleu : VV). Image communiquée par le Centre canadien de télédétection.
Relation avec la fréquence
La possibilité d'extraire des données biophysiques des données RSO dépend fortement de la fréquence. Les micro-ondes à basses fréquences, comme celles des bandes L (1,0 à 2,0 GHz) et P (0,3 à 1,0 GHz) pénètrent mieux le couvert et interagissent davantage avec ses éléments structuraux (feuilles, branches, troncs). Les ondes à plus haute fréquence (bande C : de 3,8 à 7,5 GHz) interagissent surtout avec la partie supérieure du couvert. Les capteurs à haute fréquence pourraient avoir une utilité limitée, pour la mesure de la variation de la biomasse sèche, alors que l'on prévoit que les données des bandes L et P pourront servir à distinguer les variations de la biomasse.
9.3.2 Dépendance selon la polarisation
Le Toan et coll., 1992 ![]() ont montré que la gamme dynamique de la rétrodiffusion d'un peuplement uniforme de pins dépendait fortement de la polarisation. En effet, ils ont obtenu dans la bande P des gammes dynamiques de 15 dB, 11 dB et 4,8 dB pour les canaux HV, HH et VV, respectivement. Les gammes dynamiques dans la bande L sont réduites : 8,6 dB, 5,3 dB et 4,6 dB pour les canaux HV, HH et VV, respectivement. Dans la bande C, la gamme dynamique est encore plus étroite, ils ont obtenu 4 dB, 2 dB et 2 dB pour les canaux HV, HH et VV, respectivement.
ont montré que la gamme dynamique de la rétrodiffusion d'un peuplement uniforme de pins dépendait fortement de la polarisation. En effet, ils ont obtenu dans la bande P des gammes dynamiques de 15 dB, 11 dB et 4,8 dB pour les canaux HV, HH et VV, respectivement. Les gammes dynamiques dans la bande L sont réduites : 8,6 dB, 5,3 dB et 4,6 dB pour les canaux HV, HH et VV, respectivement. Dans la bande C, la gamme dynamique est encore plus étroite, ils ont obtenu 4 dB, 2 dB et 2 dB pour les canaux HV, HH et VV, respectivement.
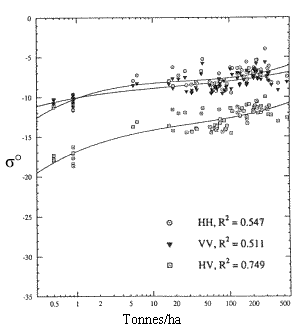
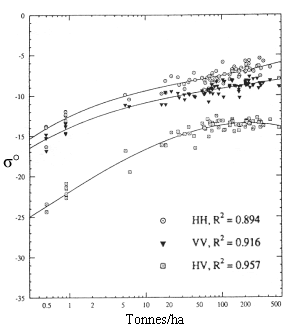
Comme le montre le travail de Dobson et coll. ![]() [voir figure 9.27], on estime que l'utilisation de la polarisation HV aux grandes longueurs d'onde permettra de caractériser plus efficacement la biomasse forestière.
[voir figure 9.27], on estime que l'utilisation de la polarisation HV aux grandes longueurs d'onde permettra de caractériser plus efficacement la biomasse forestière.
Bande C
Bande L
Figure 9-27. Graphique semi-logarithmique du signal rétrodiffusé dans les bandes C et L et étalonné, en fonction de la biomasse totale en tonnes par hectare de pin maritime et de pin à encens au dessus du sol. (Tiré de ![]() ).
).
Paramètres polarimétriques
9.3.3.1 Signatures copolarisées
Les données polarimétriques pourraient fournir des informations uniques sur le couvert forestier, notamment l'effet de l'architecture du couvert sur les mécanismes de rétrodiffusion. Les mécanismes de diffusion sont : la rétrodiffusion directe des branches (réflexion unique ou diffusion dans le volume), la rétrodiffusion des troncs (réflexion unique), la rétrodiffusion des branches et du sol (double réflexion), la rétrodiffusion des troncs et des branches (double réflexion) et la rétrodiffusion directe du sol (diffusion à la surface). Les contributions relatives de chacun dépendent de la nature du couvert et des paramètres d'imagerie, comme l'angle d'incidence et la fréquence. En fonction de la fréquence, les éléments du couvert (les feuilles et les ramilles) pourraient jouer un rôle important dans les interactions menant à la diffusion et l'atténuation. On s'attend à ce que l'analyse des données polarimétriques puisse aider à comprendre la nature de la diffusion dans le couvert.
Nous montrons à titre d'exemple la simulation de la signature copolarisée d'une forêt caducifoliée, dans la bande L. On peut y voir que la rétrodiffusion d'une couverture forestière dense dans la polarisation HH est similaire à celle de la polarisation VV, quoique moins intense. Ceci suggère que le mécanisme dominant est la diffusion multiple par les branches, auquel s'ajoute une faible composante de double diffusion (figure 9-28A). La hauteur du socle révèle que la contribution à la rétrodiffusion des échos non polarisés est importante, ce qui indique une diffusion multiple. ![]() .
.
La modélisation du même couvert avec une densité de branche dix fois moindre (figure 9-28B) produit une signature de polarisation caractéristique d'un réflecteur dièdre (soit une double réflexion), ce qui indique que dans cette forêt moins branchue, le mécanisme dominant est la diffusion tronc-sol. La plus faible densité de branche se traduit par un socle beaucoup moins haut, puisque la composante non polarisée est moins intense, ce qui indique que la diffusion dans le volume est moins intense.
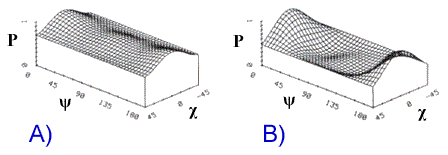
P = puissance normalisée
Figure 9-28. En A, signature copolarisée dans la bande L d'une forêt de feuillus et en B, signature copolarisée d'une autre forêt de feuillus dont la densité de branche est dix fois moindre. (Tiré de ![]() ).
).
La figure 9-29 montre les signatures copolarisées modélisées pour une région boisée et une région déboisée (coupe blanche). Le socle de la région déboisée est plus bas, ce qui indique que le signal rétrodiffusé par la région coupée à blanc est moins dépolarisé, ce qui s'explique par la plus grande intensité des échos directs du sol et l'importance moindre de diffusion dans le volume. L'intensité plus forte de la rétrodiffusion pour le cas VV, par rapport au cas HH, témoigne de la réflexion à la surface.
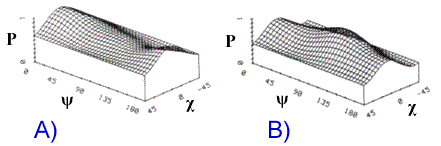
P = puissance normalisée
Figure 9-29. Signatures copolarisées A) d'une région boisée et B) d'une région déboisée. (Tiré de ![]() ).
).
9.3.3.2 Différence de phase polarisée
Les différences de phase copolarisées peuvent contribuer à la compréhension des mécanismes de diffusion d'une cible donnée. En général, les diffuseurs à réflexion unique produisent une différence de phase proche de 0°, alors que les diffuseurs idéaux à double réflexions engendrent une différence de phase de ![]() 180°. L'exemple qui suit est tiré des travaux de Le Toan et coll.
180°. L'exemple qui suit est tiré des travaux de Le Toan et coll.![]() qui ont corrélé, pour un couvert forestier, la différence de phase copolarisée avec l'âge du peuplement (figure 9-30) et, à un degré moindre, avec la hauteur de la forêt et la biomasse contenue dans les troncs. Dans ce cas, la différence de phase copolarisée moyenne d'une zone déboisée (
qui ont corrélé, pour un couvert forestier, la différence de phase copolarisée avec l'âge du peuplement (figure 9-30) et, à un degré moindre, avec la hauteur de la forêt et la biomasse contenue dans les troncs. Dans ce cas, la différence de phase copolarisée moyenne d'une zone déboisée (![]() = 6,8°) indique que la rétrodiffusion provient surtout de la diffusion de surface sur le sol. La valeur moyenne d'un peuplement forestier adulte est très supérieure (environ 66°) et son écart-type est plus élevé (90°) que celui d'une zone déboisée (70°). C'est un résultat de la diffusion dans le volume.
= 6,8°) indique que la rétrodiffusion provient surtout de la diffusion de surface sur le sol. La valeur moyenne d'un peuplement forestier adulte est très supérieure (environ 66°) et son écart-type est plus élevé (90°) que celui d'une zone déboisée (70°). C'est un résultat de la diffusion dans le volume.
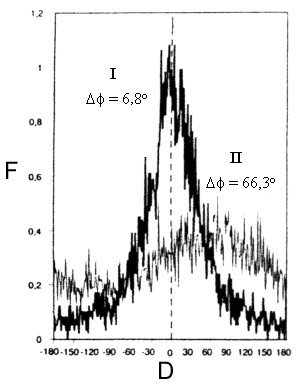
F = fréquence (%), D = différence de phase polarisée (degrés)
Figure 9-30. Histogramme des différences de phase copolarisées dans la bande P entre les polarisations HH et VV (![]() ) pour une zone déboisée (I) et une plantation forestière de pins, âgée de 46 ans (II)). (Tiré de
) pour une zone déboisée (I) et une plantation forestière de pins, âgée de 46 ans (II)). (Tiré de ![]() ).
).
Détails de la page
- Date de modification :